Journal
of the Trinidad & Tobago Field Naturalists' Club, 1975
SOME FIELD OBSERVATIONS ON THE BACHACS
OF TRINIDAD
by D. J. Peregrine
(Department of Applied Zoology,
University College of North Wales).
INTRODUCTION
Since 1969, the Overseas
Development Administration of the British Government has financed research
on leaf-cutting ants (bachacs) with the aim of devising efficient control
methods for those species which are considered pests. Several scientists
in Britain are involved with the project and at least one person is based
in South America to carry out most of the field work. Up until very recently,
the field work on this project has been done in Trinidad and this together
with the laboratory studies done in the U.K., means that much of our understanding
of these insects is based on those species occuring on this island.
Before attempting to control
a pest in the field, one must be thoroughly familar with its biology, including
its ecology, behaviour and so on. The leaf-cutting ant project is no exception.
Over the past five years we have accumulated a great deal of knowledge
as regards the biology of these ants and the purpose of this article is
to summarise for the general naturalist some of the information that has
emerged out of this and other people's work.
It would be impossible to
cover all aspects of the biology of these interesting insects in a short
article. In view of this, more attention has been paid to those behavioural
aspects which the field naturalist may observe himself.
THE SPECIES IN TRINIDAD
There are only two species
of bachac in Trinidad although several other ants are incorrectly given
this name. Most people do not distinguish between these two species but
to the entomologist and the serious naturalist the two are very different
in both structure and habits. These differences warrant the placing of
these ants in two separate genera. Atta cephalotes (L) (Fig. la-f) or the
forest bachac as it is sometimes called, has a shiny appearance, is reddish-brown
in colour and has long slender legs. This species is not confined to Trinidad
for its distribution extends from Central America to Brazil. The majority
of the nests are to be found in natural or semi-natural rain forest. Acromyrmex
octospinosus (Reich.) (Fig. lg-1) is a more ubiquitous species. The workers
are generally smaller than those of Atta and there is no soldier caste.
These ants vary in colour from brown to almost black and the legs are short
compared with those of the previous species. It nests in various habitats
including walls, under stones, under fallen tree trunks and in the ground
itself.
LIFE CYCLE
Bachacs, like many other
ants, have regular flights of winged males and females (Fig. 1) from the
parent colonies. In Trinidad, Atta cephalotes and Acromyrmex octospinosus
fly at about the same time with the former species reaching peak activity
in July or August and the latter in May or June. The winged males and females
of Atta (Fig. 1. d, e, f) are large, conspicuous insects about one inch
(2.54 cm) in length. Those of Acromyrmex are smaller by comparison (Fig.
1. j, k, 1) measuring about half an inch (1.27 cm) in total length. In
both types the sexes can be told apart by the size of the head. In the
males the width of the head capsule at its hind margin is never greater
than half the width of the thorax, whereas in the females the head is as
wide as or slightly less wide than the thorax.
A single female may copulate
with several males either from a single nest or from different nests. In
this way she builds up a store of sperm that will have to last her up to
20 years. Having mated, the young queen breaks off her wings by pushing
them with her legs and proceeds to dig into the soil. She excavates a vertical
tunnel and when this is a few inches deep (about 1 foot in Atta,) she proceeds
to enlarge it to form a small chamber slightly to one side of the main
shaft (Fig. 2). When the excavation of the nest is complete the queen spits
out a small wad of fungus from her mouth. This fungus she has collected
before leaving the parental nest. As the fungus grows, the young queen
periodically leaves the nest and searches for fresh leaves in the immediate
vicinity. These cut leaves are then taken back into the nest and are chewed
into a sticky green pulp which is added as substrate for the developing
fungus. In Atta cephalotes at least, the first viable eggs hatch to give
small workers (Fig. 1. a) in about 70 days. These ants leave the nest on
short distance foraging excursions, returning with leaf fragments for the
rapidly expanding fungus gardens.
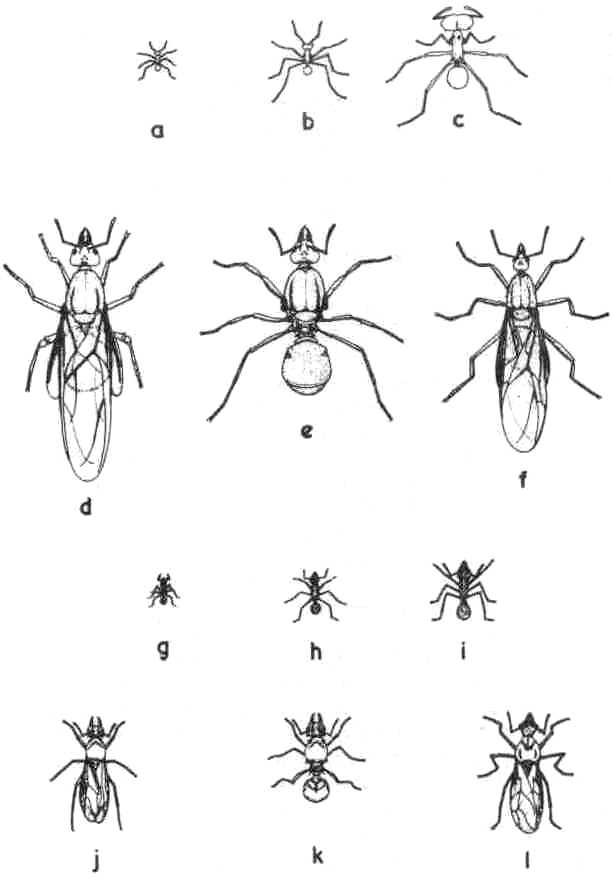
Fig 1. Various castes
of Atta cephalotes and Acromyrmex octospinosus. Atta a-f. Acromyrmex g-l.
(a), small worker (minim);
(b) medium sized worker; (c) soldier (maxim); (d) winged queen; (e) wingless
queen; (f) winged male; (g) small worker; (h) medium-sized worker; (i)
large worker; (j) winged queen; (k) wingless queen; (1) winged male.
|
DEVELOPMENT OF THE NEST
As the colony ages, so the
number of ants and the size of the nest increases. The workers excavate
more chambers to accommodate further fungus gardens. Even after 2 or 3
years, a colony of Atta cephalotes is made up of many hundreds of thousands
of individuals. Some records suggest that a 6-year old nest could contain
anything up to 5 million individuals. Colonies of Acromyrmex never reach
this size and a nest of 100,000 individuals would be considered a large
one. The smaller size of Acromyrmex nests is probably one reason why the
species is to be found nesting in many different types of habitat.
Whilst the colony is expending
obvious changes are taking place in the general appearance of the nest.
In Atta cephalotes for example, when the new queen first excavates the
primary chamber she forms a small crater with the discarded soil (Fig.
2). The crater then becomes a turret or chimney by the activity of the
first brood of workers about 70 days later (Fig. 3). The nest remains in
this condition for the first year of its life and thereafter reverts again
to a crater shape of increasing size. As the nest expends more and more
craters appear on the surface. A colony of Atta cephalotes probably starts
producing winged sexual forms after 2 or 3 years and therefore can be considered
mature at this stage, although over the next few years the nest will increase
greatly in size.
A mature nest will contain
many fungus gardens, varying from the size of a tennis ball to that of
a football. Normally the fungus gardens are concealed in subterranean chambers
and can only be examined when the nest is excavated. When the fungus garden
is examined it is seen to be made up of minutely subdivided pieces of leaf,
often withered to a brown colour, and covered with fine strands of a delicate
white fungus. At certain points the fungus appears in the form of compact
clusters, the 'staphylae', which the ants pick and feed to their larvae.
There is also a substantial
turn-over of material in these gardens. As the fungus develops so it depletes
the macerated leaves of their nutrients and eventually there is so little
fungal growth that the whole garden is broken up and discarded in the deeper
parts of the nest.
Another conspicuous activity
of the mature nest is that of soil disposal by the workers. As the nest
grows, more and more chambers have to be excavated to accommodate the increasing
number of fungus gardens. The soil from these areas is carried out onto
the surface of the nest and dumped in a selected spot.
FORAGING BEHAVIOUR
As the nest increases in
size, foraging becomes more organised. Foraging behaviour reaches its peak
of development in the genus Atta presumably in response to the large complex
social system operating here. Atta cephalotes for example builds well defined
trails about six inches wide and often over 100 yards long. Although a
large nest of Acromyrmex octospinosus has long trails leading from it,
they are never as conspicuous as those produced by Atta.
Recent work carried out in
Trinidad (Lewis et al, 1974b) has shown that both Atta and Acromyrmex have
well defined foraging cycles. Using automatic ant counters, Lewis and his
colleagues was able to record continuously over a period of several
months, the numbers of ants leaving the nest on foraging missions at different
times of the day. They found that Atta cephalotes is a predominately nocturnal
forager. Foraging commences regularly at about 19.00 hours, reaches a peak
of activity at about 21.00 hours and then subsides by about 06.00 hours.
However, not all the foraging follows this pattern. Less frequently the
ants of some nests forage during the day. Again the regularity of the rhythm
is striking. The workers in such a nest would start foraging in earnest
about 09.30 hours, build up to a maximum about 13.00 hours and foraging
would cease by 19.00 hours.
Different foraging trails
leading into the same nest would sometimes be out of phase, that is to
say that whilst the ants on one trail were nocturnal, the ants on the other
trail were diurnal. The situation is further complicated by the fact that
on certain occasions the ants on a particular trail would change from being
nocturnal to being diurnal or vice versa. This change over in 'shift work'
does not occur suddenly but the regular rhythmicity falters for about two
days before the new rhyhtm is established. The reasons for these peculiar
but interesting cycles of behaviour are not at all clear. Lewis et al.,
(1974a) have examined a variety of environmental factors such as temperature,
light intensity and rainfall in an attempt to correlate these with foraging
patterns but no obvious correlations emerged from these investigations.
They have suggested that it may be the nutritional requirements of the
brood and/or fungus gardens which determine whether foraging occurs at
night or during the day, since it is well established that the nutrient
content of leaves alters at different times of the 24-hour day.
In tropical rain forest with
its abundance of plant species, Atta cephalotes will cut the leaves of
many different plants. For example, Cherrett (1968) working in Guyana for
a 10-week period recorded a single colony as having cut at least 16 different
types of plant in this time. Among this range of species, some were obviously
preferred to others, and some species also present in the forest were not
attacked at all. That bachacs attack different plants is not unusual in
that many species of insects and other animals are polyphagous or 'general
herbivores'. What is strange and something appreciated by the astute naturalist
is the spatial distribution of bachac foraging. Examination of an Atta
nest in the forest, and the vegetation in an area 50 yards radius from
it, will soon reveal that the ants travel far to cut particular trees when
there are specimens of the same species closer to the nest. What then are
the reasons for this behaviour? Attempting to explain the situation, Cherrett
(1968) has put forward the following plausible theory. He believes that
the foraging system used by Atta is efficient in conserving the natural
balance of plant species in the forest. If a large Atta colony attacked
all the suitable specimens nearest it, then this sort of grazing pressure
would certainly kill many of them off. These cleared areas would probably
then be invaded by species resistant to leaf-cutting ant attack. There
is no evidence to suggest in fact that the plants nearer to the nest are
any more resistant than the ones further away.
The process of leaf cutting
is interesting and the field naturalist would be well advised to spend
some time studying it. The first thing to notice is that young foliage
is attacked in preference to old. Physical factors are partly responsible
for this. older leaves becoming tougher and therefore more resistant. Barrer
and Cherrett (1972) have also shown that chemical factors are also important
in determining which leaves on a particular plant will be attacked. It
seems that young leaves possess attractive chemicals which induce cutting,
but when the leaf gets older there is a build-up of repellent chemicals
which tend to counteract other leaf cutting stimuli.
Another feature worth noting
is that of the pattern of cutting on individual leaves. Examination of
Figure 4 shows that the cuts are not evenly distributed but that once a
cut has been made it acts as a focus for further cutting. Barrer and Cherrett
(1972) examined this phenomenon in some detail and were able to show that
a cut edge has both physical and chemical properties that make it more
likely to be cut than an intact leaf edge.
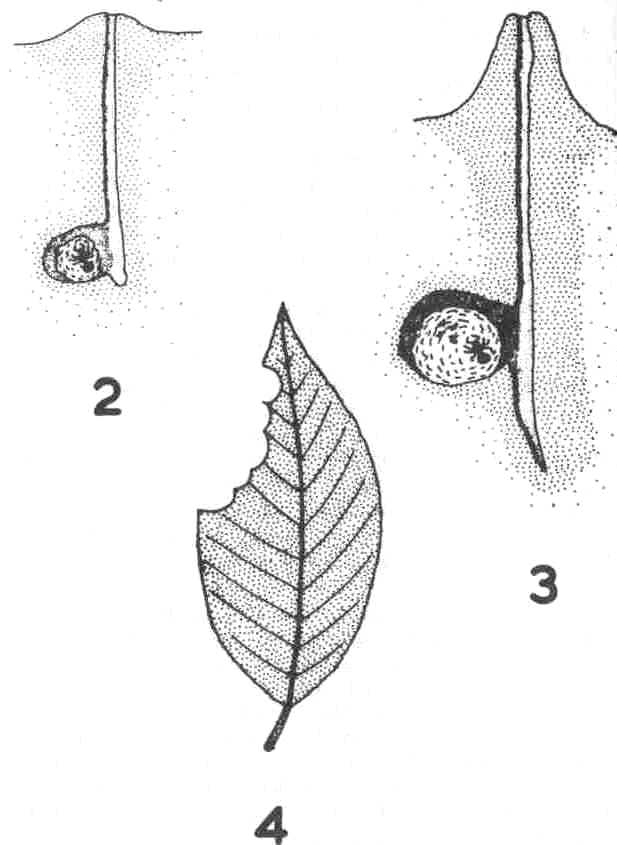
Fig. 2. Young nest of Atta
cephalotes at the small crater stage.
Fig. 3 Slightly older nest
of Atta cephalotes at the chimney stage.
Fig 4. Typical Atta cephalotes
damage to a leaf.
|
During leaf cutting the mandibles
of the worker ant are used in very much the same way as one uses a tin
opener. One mandible is inserted onto the leaf tissue and does not contribute
much to the actual cutting whilst the other mandible is pulled through
the leaf. Having completed the cut, the ant manipulates the leaf segment
into a position ready for carrying. Because the fragment weighs as much
as or even more than the ant itself, getting it into the correct position
is important. The leaf is normally carried with its centre of gravity directly
above the ant.
Close examination of a column
of Atta foragers will reveal the presence of small worker ants (minima)
on many of the leaf fragments being carried back to the nest. Usually
there is only one minim per leaf section but there can be more. What then
is the function of these 'hitchhikers'? Weber (1972) proposes that they
serve a dual function. In the first place they lick the surface of the
leaf and it has been suggested that they are removing debris and alien
organisms. The fungus cultured by the ants is a pure culture and therefore
it is important that foreign fungi are excluded. Another function of the
'hitchhiker' is that they fend off parasitic phorid flies. Some of these
flies are known to lay eggs on the adult ant in which they develop as internal
parasites. When these phorids are about, the minima workers rear up on
their middle and hind legs and fend off the flies with their fore legs,
antennae and mandibles.
OTHER ANIMALS ASSOCIATED
WITH THE ANTS
A variety of animals is associated
with the bachacs either as guests, parasites or predators. Mites, springtails
(Collembola), nematode worms and phorid flies are to be found in the nests,
whilst some amphibians and mammals are predators. The common giant toad
(Bufo marinus) in Trinidad feeds on bachacs as well as on many other ant
species. Also the silky anteater (Tamandua longicauda) takes both the species
of bachac in its specialised diet. Another interesting animal, the legless
lizard (Amphisbaena) is also quite commonly found in nests of both Atta
and Acromyrmex in Trinidad.
REFERENCES
Barrer, P. M. and Cherrett.
J. M. 1972. Some factors affecting the site and pattern of leaf-cutting
activity in Atta cephalotes (L.) (Hymenoptera: Formicidae). J. Ent. (A).,
47: 15-27.
Cherrett, J. M. 1968. The
foraging behaviour of Atta cephalotes (L) (Hymenoptera, Formicidae) I.
Foraging pattern and plant specie attacked in tropical rain forest. J.
Anim. Ecol., 37: 387-403.
Lewis, T., Pollard, G. V.
and Dibley, G. C. 1974a. Micro-environmental factors affecting diet patterns
of foraging in the leaf-cutting ant, Atta cephalotes (L). (Formicidae;
Attini). J. Anim. Ecol.,43: 143-153.
Lewis, T., Pollard, G. V.
Dibley, G. C. 1974. Rhythmic foraging in the leaf-cutting ant Atta cephalotes
(L.) (Formicidae; Attini). J. Anim. Ecol., 43: 129-141.
Weber, N. A. 1972 Gardening
ants, the Attines. American Philosophical Society, Philadelphia.